論文「人間による生態系の拡張:食料生産と科学の2045年目標」邦訳

この記事は、以下の論文 “Human augmentation of ecosystems: objectives for food production and science by 2045” をソニーコンピュータサイエンス研究所リサーチアシスタント 兼 (社)シネコカルチャー講師・太田耕作が訳したものです。

人間による生態系の拡張:食料生産と科学の2045年目標
株式会社ソニーコンピュータサイエンス研究所 舩橋 真俊
序章
食料生産の歴史は人類の最も素晴らしい功績のひとつですが、それによる環境負荷という功罪でも特徴づけられます。1万年以上前からの農業の発展と産業革命によって可能になった農業技術の進歩によって、今日では人口が75億に達しました。これらの進歩は世界規模で生態系に大きな影響を及ぼす文明や発達した社会システムをも創りだしました1。化学合成肥料や農薬の集中的な使用、そして高収量作物品種による緑の革命が世界規模で人類の多くを救いましたが2、その負荷が地球生物の第六の大量絶滅を引き起こしています3。ここで、物質資源、人間の健康・生態系の健全さという点から一次産業・食料生産の持続可能性に疑問が生じます。既に、農業における窒素・リン・炭素などの投入による生物地球化学的な流出は地球の許容量を超え、不可逆的な遺伝的多様性の損失が劇的に生物圏を変化させる可能性があります4。また医療費は先進国・発展途上国どちらにおいても最も経済にのしかかる費用のひとつになりました5。これらをひっくるめた問題は、慣行の食料生産システムと、食・環境・健康のトリレンマ(三つ巴構造)に繋がっています6。持続可能な方向へ再び戻るには、現在の食料システム(生産・流通・消費)のドラスティックで根本的な転換が必要です7。
食料生産と消費に関する時空間的スケールは、グローバルな環境から健康に関わる分子機能までに渡ります。図1は科学技術の領域とそれの環境へ与える影響の関係の概略を示しています。食料に関わる生物学的研究のほとんどは、僅かな数のモデル生物あるいは組織におけるin vivo試験あるいはin vitro試験という高度にコントロールされた実験室環境で行われるため、及ぶ領域がある程度制限されます。これはフードサイエンス(食品科学)の方法論的制限への批判ではなく、むしろ食料生産においてそういった要素を注意深く考慮することの重要性を言いたいのです。生産現場と研究室間のフィードバックの評価が、広い文脈ではもっと重要になってきます。
実際の食料生産のスケールはモノカルチャー栽培(in cultura)から自然生態系での狩猟採集(in natura8)にまで渡ります。例えば、農学はモノカルチャー栽培での収量の最適化に重きを置いており(すなわちin cultura)、生態学は主に生態系(in natura)の自然な振る舞いを色々な撹乱段階で調べます。生態学において生産量の最適化は、in culturaにおける個体(植物・動物など)の最適化としての生理学的最適化と、in naturaにおける複数の種が他種と相互作用する中で自然発生的に自己組織化される生態学的最適化とに定義されます9。現代社会は人間活動に影響された自然環境の強い制限の下にあり(in cultura)、ほとんどの医学研究もここで行われています。人口増加の圧力による自然生態系の損失は、人間が農地や生活圏においてin naturaをin culturaで支配してきたように、農業の歴史上ずっと続いています。人間活動によるグローバルな生物地球化学的な循環の変化によって、今の地質時代は人新世(Anthropocene epoch)と呼ばれています10。
図1は、y軸の時空間スケールによって生じる事柄をx軸の科学技術の複雑性の関数で表しています。都市と農地の開発における既存のシナリオは砂漠化などのステートシフト(状態遷移)を最終的にもたらしますが1、革新的な未来の食料生産の科学技術は自動農業や植物工場(似たものとしてin vivo試験9やin vitro培養肉11など)のような、環境条件をコントロールすることに集中する傾向にあります。こうした、新たに生じつつある代替案は高度にコントロールされ閉鎖された環境(cGMP実験室など)に基づいていて、その極致には宇宙船内環境があります。
クローズドシステムを使用した研究(例:実験室)に基づいて、これまでの科学は開かれた環境にその知見を(多様な負荷をかけつつ)応用してきました(図1の紫色の破線の中)。ここからは、よりよい枠組みとしてオープンシステムサイエンスと呼ばれる考え方が必要です(図1赤色の破線の中)。オープンシステムサイエンスは問題解決時にまわりのシステムとの関係を重視し、広範囲に渡りコンピュータ技術を問題の領域・構造・機能の定義に用いることで、タイミングよく効率的に、限られた観測条件の中でそれをコントロールすることができます。与えられた気候条件の中でin natura生態系のマネジメントをすることは、衛星によるリモートセンシング、クラウドコンピューティング、人工知能、IoTのようなICTによるサポートの開発とオープンシステムサイエンスの導入によって拡張された生物多様性の実現を可能にするポテンシャルを秘めています13-16(図1の右上)。これは慣行農法にICTを導入することに基づく「スマートアグリカルチャー」とは質的に異なっており、その拡張は正味の生態系機能を自然状態より高めます9。一般的に生態学的最適化とは、進化的に発達した自己組織化能力は群集レベルでの生態系機能のある種の包括的最適化とする仮説に基づいて生態系が作り出した状態を意味します(すなわち自然に形成された生態系の多機能性naturally formed ecosystem multifunctionality17)。その植生と調和して発達した生態系の性質や、生態学的遷移の多様性と代替安定状態(alternative stable states)の存在18,19によって、同じ環境条件における異なる種構成から生じる生態系機能に差異が生じ得て、また色々なレベルの生態系サービスをもたらします。これは、種多様性を人工的に操作することによって生態学的最適化は自然状態に比べて拡張可能であることを示唆します。
図1のY次元(紫と赤の矢印)は生物多様性への人間の影響を正(上方向の赤)・負(下方向の紫)で表しています。効果的な緩和シナリオは環境負荷を自然の許容量を超えないようにすべきであり、正味で損失を相殺しプラスになるように高度にin naturaで自己組織化された生態系の拡張が必要です。しばしば、生物多様性オフセットやカーボンニュートラルなどに関する主張はベースラインをその許容量ではなく現状の負荷としてしまっています。例えば、生物多様性が高いといわれる農地も必ずしも自然が保護されるレベルより高くなく、そもそも初期に農地に転換した時点で損なわれています。このことは年間の温室効果ガス排出の6分の1を占め21、先述の大量絶滅の主要な原因となっています(気候変動による影響よりも大きい22のです)。
環境負荷と可能な緩和・回復の政策について理解するには、それらを含む複雑系に関して多段階における統合的な枠組みが必要です。現状の負荷が社会的・生態学的ステートシフトを今世紀半ばに引き起こすと推定されるため、マテリアルサイクルや生物多様性のオフセット、回復について適切に評価する方法をつくることが根本の課題となります。これについては下記のことを必要とします。未来に向けた持続可能な食料生産の強化(森林保護のために菜食主体への文化・食料システムの転換を含む)24,25、不適切な生産・流通の改善26、都市農業における地産地消の推進27、昆虫食のような環境配慮と栄養学的に価値の高い代替食の探索28、資源の少ない農家のための小規模農業の多様な仕立て29、生態系の機能的キャパシティに基づいた漁業の管理30などです。さらに、そういった手段には人口増加と社会的不平等に対応するプラスの影響を得るために、緩和策を超えた環境と健康へのリスクの解決も組み込まれるべきです6,9,20。
気候変動の観点では、統合的なアプローチが重要です。農業、林業、その他の食に関連する土地利用は人間による温室効果ガス排出量の25%を占めています31。気候変動に最も脆弱な国は、社会の腐敗32や飢餓33、特に沿岸地域の生物多様性ホットスポットを有しています34。こうした国での食料生産は主に小規模家族経営の農家によって担われており、これは世界の農地の87%を占めています35。また、小中規模の農地が最大で世界全体の77%の主要な商品作物を生産しています36。
明らかに、これらの問題は現状の食料生産の最適化技術(通常通りのシナリオや先進国の研究開発投資の方針に基づいた精密農業precision agricultureなど)の及ぶ範囲の外にあります。例えば、アメリカでは大規模農場における精密農業の採用はわずか1〜3%の正味の利益しかありませんでした37。ヨーロッパでは、少なくとも調査されたケースやシナリオにおいて、インプットの部分的削減にも関わらず、実装の費用がかさみ全く利益を生じさせることができませんでした38。これらはそもそも質的に環境に負の影響を与えることが前提のものであり、モノカルチャーを効率化して広げていく方法では、世界人口の増加や見込まれる生態系崩壊に対して対抗できません39。遺伝子組み換え作物生産などの先進的な農業技術でさえ、必要なレギュレーション・サービス(これは天文学的数の遺伝資源を持つ自然植生によってのみ涵養されてきました)をもたらす安全で十分な生物多様性を保証できません。むしろ開放された場における遺伝子組換え作物(GMO)は、野生種との交雑や種間の遺伝子伝播(ゲノム進化の一般的なメカニズム)による遺伝子多様性の均一化のリスクを伴います40。
科学的知見の世界的統合によって、過去25年以上にわたる地球システムの崩壊への道筋に警告が出される結果になりました41。もし持続可能な食料生産という分野が存在するのならば、それは研究・政策決定・生産・流通・消費・リサイクル・健康効果すべてのバリューチェーンを通した人間活動によって生物多様性が調整あるいは促進されるという最終ゴールに向かって、in vitro・in vivo・in cultura・in naturaの多段階相互作用における相乗効果やトレードオフをすべて組み込んだものであるべきです。人間以外の自然と共にこの人新世の生物圏を持続させる社会をデザインすることが重要です42。農学と生態学の統合によって、フードサイエンスは人類と自然両者の健康と豊かさを提唱する統合的生命科学として、生態学的な効果・利益を長期に渡って得る食料生産に対して中心的な役割を果たすことができます。
図1
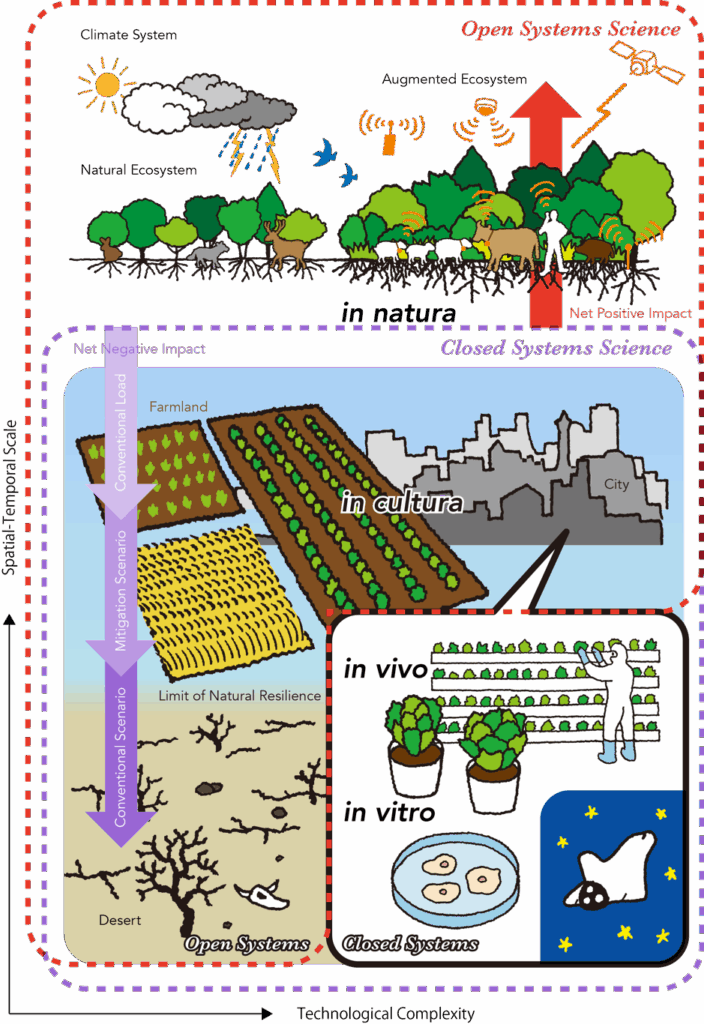 図1
図1
要素主義の問題と真の因果関係
フードサイエンスにおける現在の国際的なイニシアティブは、成分分析とそれが人間の健康に与える影響に焦点を当てる傾向にあります(例:参考文献43)。この、研究室から人体やフィールドへの外挿法は農学・医学に共通であり、科学における要素主義的方法論の基礎を形成しています。しかしながら、現状のアプローチを量的に拡大することは必ずしも問題解決への道を保証しません。奇妙なことに、我々が部分的な科学的根拠(エビデンス)を積み重ねれば積み重ねるほど、より多くの隠れた変数が無視されていきます。このことは適切な統合ができなかった無数の矛盾する部分的エビデンスから構成されている可能性があり、あるいはグローバルなコンセンサスがそういった見えないバイアスを考慮できていないままであるという結果に繋がっていることが考えられます。「木を見て森を見ず」の科学者が、複雑なシステムとして生きている生命を扱う分野には多くなる危険性があります。このアナロジーでは、木は個々のオーミクス研究で、森は人間や生態系の統合的な健康を意味します。典型的な例として言えば悪化する製薬産業の研究開発効率が挙げられます44。
率直に言えば、統計テストしか用いないハイスループットの探求的な研究は方法論的な落とし穴を併せ持ちます。研究後の主張が本当に正しい可能性は著しく低いですし、それは専門家の意見によってサポートされた経験的エビデンスのある分野においても、なのです45。p値の有意度α=0.05というのは単に第一種過誤(偽陽性)の閾値を定義するのみで、これはつまりいかなる帰無仮説も20回の実験をすることで棄却され得ることを意味します。結果として出版された論文の95%が偽陽性の可能性があり、これは実際の臨床における薬剤の試験の非再現性の割合(例えばがん研究の分野では、89%の画期的発見が再現できなかったようです46)とそれほど変わりません。
一方で、真に有意な効果でも検定力が低い場合には必ずしも検知できません。統計の結果だけでなくオッカムの剃刀(すなわち、現象を説明するのに必要十分な最も単純な考え方)に耐え得る真の関係が、生体システムの複雑性という文脈に結びつけられるべきです。生化学的経路と遺伝子相互作用の一般的性質として、生理学的な応答が多くの要素(過剰になっても不足しても致命的であり中程度の量で非線形的な正の効果をもたらす生理活性物質や、ファジィ論理のように存在量によって生物学的反応にゆらぎをもつ複数の化合物の相互作用や、下流の受容体において生理学的効果を逆転させ得るダウンレギュレーションの起動や、生物学的研究でほぼ未導入である3つ以上の変数が成す高次相関47)の複合的な効果として現れます。システムレベルの性質として、これらの相互作用は生物個体の生存に部分的に必要な条件でありますが、必ずしも要素的な研究では検知できません(真の必要条件の一部を形成する要素を誤って不採用にしてしまう可能性)。その上、生物学的システムのロバストネスというのは必ずしも生存に関係ないが環境への適応度に有意な違いを見せる周辺部の効果も意味します(必要条件ではないものを誤って採用してしまう可能性)。
ここでキーとなる課題は、観測された相関から、偶発的な関連や偽相関ではない真の因果を区別することです。有意な相関は隠れた因果関係が見つかるのを妨害したりそのノイズとしてはたらいていたりすることがあります。統計的分析は普通、偽相関と真の因果関係を区別せず、どっちつかずの結果をもたらすことがあります。長期の健康保護について微量成分が重要であるという話に関して言うと、抗酸化物質はそのような複雑な複合効果をもたらす典型的な例です。細胞レベルで有意な抗酸化作用をもたらすというフラボノイドのin vitroとin vivoにおけるエビデンスがあり48、平均寿命が長い地域での食文化にある自然の食品に多く含まれているということもありますが49、それでいてβカロテンやビタミンA、ビタミンEのような抗酸化サプリメントは大規模な臨床試験で健康あるいは安定状態の被験者の致死率の上昇をもたらすという結果50になっています。こうした不協和は食品の成分や生理化学的性質をシステム的な健康の必要条件だと盲目的に決めてしまっていることによります。もし偽相関と真の因果関係の有意性がスケールの違いによって変わるとしたらどうでしょうか。食品成分と代謝マーカーは確かに相関していますが、その(偽)相関に外からはたらいている潜在的な変数が存在しているかもしれません。現象Aと現象Bの統計的相関は必ずしも因果関係を意味せず、潜在変数Cが存在していて同時にAとBに影響しているという真の因果関係があるかもしれないという疑問が生じます。典型的な例は、(消費者の健康に関連した)植物の代謝状態や他の食品の成分に影響する栽培(生産)条件です9。健康状態と強く相関する栄養学的要素は、その観測の外に存在する他の真の要因の単なる副産物であるか、あるいはその相関をもたらす真の要因の単なる一部である可能性があります。このことが、フードサイエンスにおいて終わりなき妥当性評価・検証実験をせざるを得なくさせます。成分重視で摂取試験を行うような視点から外れ、食品全体としての健康効果を評価しようとした時に、真の原因となっている要素を探し当てることは格段に難しくなります。
そのようなエビデンス構築における複雑性は、実験条件やデータの制限を超えて統合的視点を得ることについて「悪魔の証明」的な重荷を作り出します。潜在的に関係している変数を広範囲に含むような十分に包括的な設定なしに、真の要因を区別するようなテストはできません。異なる領域の変数が互いに影響したり絡まったりしているような現実の状況では、異なる分野の個々の努力では、(そもそも方法論的に)統合された枠組みを発見することができません。こうした困難を緩和し統合的な多段階スケールのモデルを構築するため、「リスクファクター(危険因子)」や「有益成分」のような医療・食の科学の専門用語が統合されるべきです。それが図2(a1-a3)の「暗礁モデル(hidden reef model)」で、そうした用語を潜在変数として表しています。
図2
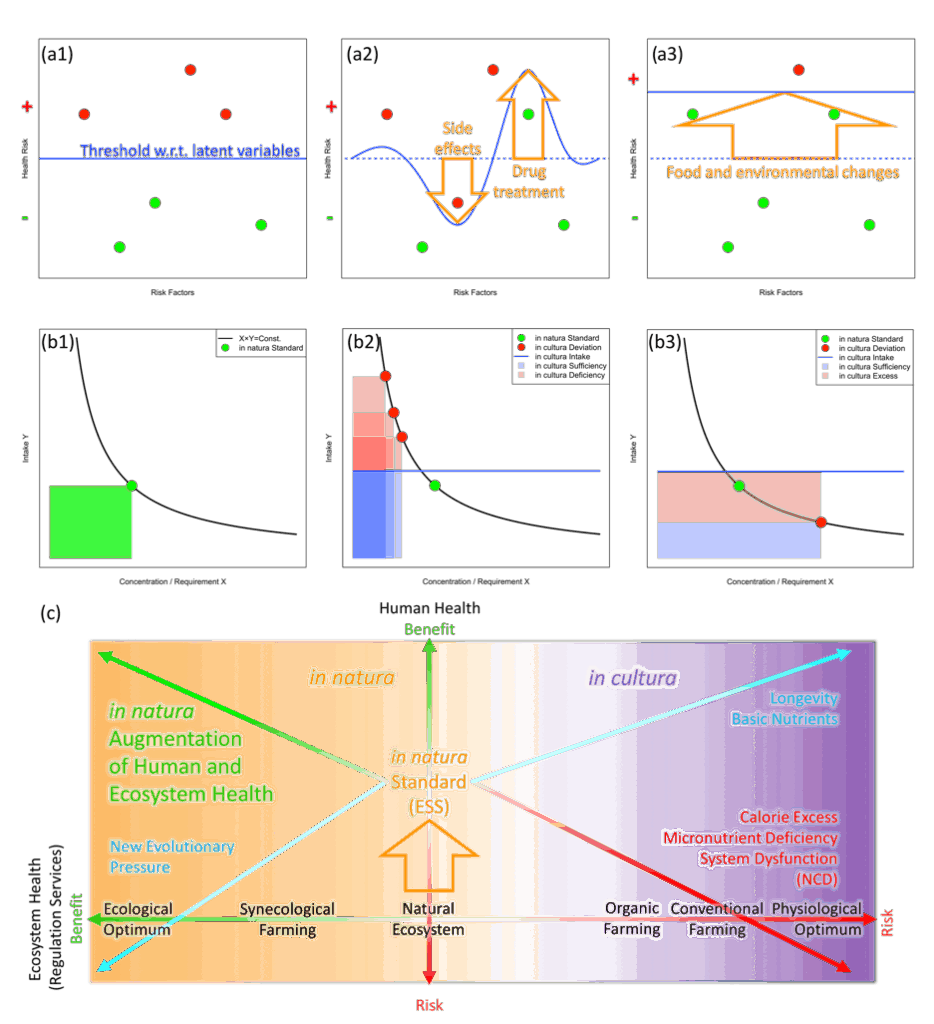
暗礁モデルにおいて、システムバイオロジーの見地から、様々なフィードバック機構をもつ複雑な生命体は、加えられた要素に対しての生物学的応答というコンテクストを提供する内部の代謝状態のような記憶構造を持っています。この内部状態は、遺伝子の変異や環境要因、社会的地位、その他のランダムなメカニズム(普通は測定することや基準をつくることが難しいものです)によって、影響を受けます51,52。図2の(a1)では、個体の健康について観測可能な要素は「暗礁」として表現され、一部が「海水面」(青線)に沈んでいます。海水面の位置は、観測不能であり潜在的な変数(環境条件や内部の代謝状態)の効果を統合した健康効果の閾値を表しています。海水面はin vitro、in vivo、in cultura、in naturaのそれぞれで変わる観測条件を表現します。海水面から出た暗礁(赤)はリスクを表し、沈んでいる暗礁(緑)は利益(健康への寄与)を表します。(a2)は薬剤投与のケースを表しています。薬剤の効果で海水面を部分的に動かし、1つのリスクファクターを減らしていますが副作用で別のリスクファクターを生じさせています。(a3)は海水面そのものを上昇させるような、適切な処置を表しています。それは環境を変えることや、あるいは暗礁を下に押し下げるように個々の特性を変化させることです。そうすることでほとんどの暗礁が水中(リスクの閾値下)に沈みます。進化的安定状態(ESS: Evolutionary Stable State)は遺伝的・代謝的なネットワークのロバストネスを形成しますが、これが上記のin naturaの食・環境条件における「隠れた暗礁」の状態に値すると考えられます53。暗礁と海水面の例はsupplementary materialの5に示してあります。
この形式化は、遺伝力の欠損問題54のような、遺伝病理学の古典的問題を食事パターンのような環境要因との関係に当てはめることができます。同じ遺伝子をもつ実験動物の表現型の多様性51(実験における背景の効果として働く)も表します。普通は、海水面は個々の研究において暗黙のうちに平面的閾値として設定されます。しかし、異なる試験の関係をマッピングするには潜在変数(異なる実験系間のインターフェースとしての可変的な海水面)を参照することが必要です。最近出てきている、長期にわたる健康や幸福の改善を狙ったオーミクス研究を適切に解釈する条件は、分子から環境までの多段階の潜在変数を統合することであり、これが現実に必須です。
栄養素の欠乏あるいは過剰による健康問題は対処が簡単ですが、近年出てきている非感染性の疾患は典型的に遺伝子レベルから環境レベル(食べ物や生活習慣)の多段階の変数に影響されていて、これがある特定の要素の欠陥ではなくてシステム全体としての問題を引き起こしています5。これらの変数は食、内分泌、神経系、腸内微生物叢、周囲の生態系、そして現在我々が部分的理解しかできていない代謝経路に渡って様々なフィードバック作用を成しています(例:参考文献55)。
複雑な生体のシステム欠陥を適切に解決するには、すべての変数について同時かつ極めて正確な計測が必要ですが、十分なダイナミクスを適切なスケールを選択することでのモデリングによっても実現できます。言い換えると、科学的方法論は還元主義に強く基づいていますが、我々がある現象をまとめる(還元する)要素はそのシステムの多様なスケールの中で選ぶことができるということです。相互作用のシステムの異なるスケールの中から、適切にそれを説明できる変数を選ぶことで、単一のオーミクス研究では対応するのが難しい潜在変数の重要な効果を統合できるはずです。複雑系の問題を前にして還元主義の挑戦と発展は次の三段階にまとめられます。要素還元主義、限られた環境でのシステム理論、そして開放された複雑系のリアルタイムでのマネジメントです(supplementary material 6を参照)。
in natura統合生命科学へのインターフェース
in naturaにおける進化的スケールが我々の遺伝的、代謝的プロフィールを形成したということ53について考えてみましょう。自然選択は適度に変動する環境のもとで遺伝的組成の長期的適応という、進化的安定状態(ESS: Evolutionary Stable State)の実現を理論的にもたらしたと考えられます56。現生人類は基本的な代謝システムを、最近になって農耕が始まるまでの狩猟採集生活に数百万年かけて適応させてきて、解剖学的にはおよそ30万年前まで遡ることができると考えられます57。農業とそれによる人新世の環境の変化による生活スタイルと食の変化は、様々な科学の分野が分かれている現在の食料生産の複雑性(図1)を生み出しました。
食-環境-健康のトリレンマを解決に向けて関連する分野を統合するため、食の持続可能な生産を支える科学はインターフェースとして始原にある原因を組み込むべきです。その原因とは、農業によるin naturaからin culturaへの食の生産条件の急激な転換です。そして次に、その巨視的な条件がどうやって環境や健康において観られるような細かい問題(各分野が必ずしも他の分野を参照せずに奮闘しています)をもたらしたのか、考えるべきです。
図2(b1-b3)は、暗礁モデル(a1-a3)との関係の中で、in naturaとin culturaの区別がどうやって食-環境-健康のトリレンマを説明するインターフェースになっているのか、進化的安定状態(ESS)との対応による食の変数の収支モデルで示しています。(b1-b3)ではx軸があるひとつの食品あたりの量を生理学的あるいは環境的に必要とされる量(栄養成分、生産による環境への効果など)で割ったものを表します。栄養素の場合は、x軸の値はあるひとつの食品の重さあたりの栄養素量に対応します。これは食のガイドライン58で普通言われるような栄養素密度とエネルギー密度の掛け算とも一致します。y軸は実際の食の摂取量、すなわち食品の重量などを表します。(b1)では食の変数(x軸)とその摂取量(y軸)の緑の円に標準化され、in naturaのESS(自然状態において栄養素需要曲線 x×y = 一定 の標準点)を表現しています。in cultura環境に当てはめる(b2-b3)と、食品変数(x軸)が変化することで赤の円にシフトし、個体の生理学的な要求に影響を与えます(青線の緑の円を超えた上方への移動)。赤い円における食品の必要量(y軸)は緑の円における栄養の需要を満たすものであり、青線がそれによる個体の応答を表しています。
栄養失調の典型的な例として、微量栄養素欠乏(b2における左に動いた赤い丸)の高カロリーな食品は過食(青い線の上昇)を引き起こします。b2とb3で赤く塗られたエリアは微量栄養素欠乏とカロリー超過をそれぞれ表します。この青線の上昇は2つの意味があります。1つ目は、動物は目の前に多くの食べ物があると必要量よりも多く食べてしまうということで、2つ目は、食品の微量成分の欠乏によって栄養要求を満たすために摂取量を増大させてしまうということです。生態学的な文脈では人間の食性の進化もエネルギー密度が高く口当たりのよい食を選んできたことを支持しています59。健康リスクに関しての食品成分分析の例として、赤い円のx軸とy軸の値は先述の「暗礁」に対応し、緑の円のESSから離れた青線が「海水面」に相当します。ここでのキーとなるアイデアはある栄養について同じx軸y軸の値の意味が個々人によって変わるということです。青線の位置によって適切な量や過不足が決まります。
農業による環境負荷の例で考えると、(b2)と(b3)の赤い円で、in cultura sufficiency、deficiency、excessは必要な負荷、減少された負荷、過剰な負荷とそれぞれ読めます。(b2)での左へのシフトは狩猟採集よりも少ない負荷で多くの食料を生産できることを表し、(b3)での右へのシフトはある食品あたりの生態系破壊効果が大きいことを表します。青線の上昇はこの例では農業による収量上昇を表します。食の変数やin culturaにおける応答についてはsupplementary materialの7を参照してください。
ESSでの摂取バランスをin naturaでの生態学的最適化とすると、in culturaによるシフトは食の変数の過剰/不足になり、健康リスクや環境負荷の増大をもたらします。人間と環境の両者の健康にとっての必要十分条件は、食の一次生産にin naturaを再導入すること、すなわち拡張された生態系の多様性の中で我々の代謝をESSの状態に維持することであると言えるでしょう9。一次生産へのそうしたアプローチは生物多様性への正味のプラスの影響(net positive impact)アプローチ20と整合性がありますし、これが世界の小規模農家に導入できれば、食-環境-健康のトリレンマを解決するボトムアップの解決策になる可能性が有ります60-61。
人間と生態系の健康の関係が図2の(c)に図式化されています。農業生産の様式や結果としての健康効果・リスクを表現しています。歴史的に、生理学的最適化のモノカルチャーという方向に農業システムが発展していったことは平均寿命の上昇、基礎的な栄養状態の改善をもたらしました(右上のシアンの矢印)。同時に、非感染性疾患のリスク要因として食品内の微量成分とカロリーのバランスが悪くなったり、農地や周辺生態系のレギュレーションサービスが深刻に減衰したりしています(右下の赤い矢印)。生態学的最適化による解決策は、食料生産のために植物群集の自然発展の中に人間による生物多様性を導入することで生態系機能を高めることです。これは人間と生態系の健康に相乗効果的に貢献する可能性を秘めています(左上の緑の矢印)。各生物個体の生理学から群集の自己組織化の生態学に渡り多様なスケールで適応する過程に多様な機能の補完や相乗効果をもたらします9。この拡張シナリオ(augmentation scenario)におけるリスクは、食や生態系の変化による新しい進化的圧力が現れることです(左下のシアンの矢印)。例えば、出生率の低下や高齢化は人間開発指数(HDI)の上昇によってもたらされました62し、新たな食習慣による世代間にまたがる健康リスク63が現れたり、人間社会と保護された野生生物の衝突が高度に開発された生態系で起きたり、新たに進化した病原体が現れるかもしれません。
in natura生態系拡張で農業によるレジームシフトを阻止・逆転
我々が食料生産をリフォームすることで、人新世においてどういったことが起きてくるでしょうか。農業の普及を見てみると、この地球システムの機能に影響を与える大きな力は、天文学的・地球物理学的な力から人為起源的な力へとシフトし、これは第四紀後期の氷河期・間氷期のサイクルを脱するリスクを抱えています64。一次産業・食料生産においてin naturaダイナミクスを再獲得し強化することは、予測される世界規模の生物多様性の崩壊1などのレジームシフトや、社会システムや健康状態の悪化23への対策として必要な方法となるでしょう。
実践の経験から得られたエビデンスによって、人間活動が、環境のトレードオフを根本的に乗り越えて、生態学的最適化というin naturaの自己組織化の下で生物多様性を保護状態よりも高めることができると示されています13,60,61,65,66。食料生産におけるin natura生態系拡張は、環境への影響の質的な変化(ネガティブからポジティブへ)を伴って、最近際立った結果を残しています。それについて下記で議論します。
高い生物多様性をもつ農地や自然農法は、ポジティブな撹乱や植物種の導入によって周辺の景観を保護状態よりも生物多様性が高い状態へと変化させると知られています9。その極致としての案として協生農法プロジェクト(図2(c)のsynecological farming)が、生態学的最適化の中で、商品作物用農地の種多様性を高めることと高密度な混生状態をつくることで、in natura生態系の拡張へのエビエンスを蓄積してきています65。日本での実験では、3000㎡の土地に、名高い保護価値をもつ伝統的農地の地域的なスケール(県規模)に匹敵する作物種多様性を有することが可能であることが示されています13。周辺生態系を含む種多様性記録においては保護価値のあるIUCN絶滅危惧種が観測され、高度に組織化された土壌微生物の多様性やそれに付随した緩衝機能、高い収量、授粉作用を含むレギュレーションサービスの向上が見られました61。これは、耕起・施肥・化学物質(農薬)を、拡張された生態系機能/サービスで完全に置き換えることで、(持続する収穫圧の中で土壌の状態を損なうどころかむしろ高めて)その多種多様な産物の総合的な結果として生産性がさらに優れることを意味します。アフリカのサブサハラ、ブルキナファソにおける実験では劇的に局所的レジームシフトを逆転させ、種の構成において一次遷移の成熟した段階の植生を再構築しました60。その生産高は当国の統計において市場作物の40倍から150倍に達し、トータルでのコスト効率は10倍でした67。これらのことは、前例がなかった高度な生物多様性の向上と局所的経済の活性化を両立させることができるという、乾燥地農業の持続的開発ができる範囲を拡大させる鍵となるエビデンスと言えるでしょう。協生農法は作物多様性と収量を高める巨大なポテンシャルを秘めていますが、これは土地の開墾を最小化し、結果としてその脅威にさらされている大きな哺乳動物たちの住処を守ることにもなります68。
上記結果を総合すると、人為起源的な力によって生物多様性や植物群集の生態系機能を生態学的最適化による成長で高めることができ、これをin natura生態系拡張(in natura augmentation of ecosystems)と表現できます。生物多様性が自然状態より高いというときに、これは3つの種多様性の考え方に基づいて次のような意味を持ちます。
1、
α多様性:ある特定の生態学的遷移段階において、種の多様性を人間による作物種の導入やそれによる自然発生種の出現によって拡張することができる。
2、
β多様性:ある2つの異なる生態系(異なる遷移段階)で種多様性が異なるかどうかに対応する。これは複数の生態系を異なるα多様性でマネジメントすることで高めることができる。
3、
γ多様性:様々な遷移段階のすべての生態系における種多様性を表す。これは農地と周辺生態系の相互作用を使って拡張と保護を組み合わせることで高めることができる。
この種多様性の3段階を、種の導入やポジティブな撹乱という人間の介入によって、自然保護よりも早く狙った状態へと高めるマネジメントをすることができます。このプロセスは、導入された非侵襲的な種の貢献だけではなくて、侵襲的だとしてネガティブに見られしばしば追い払われるような優占種のポジティブな側面も含みます69。通常、ある遷移段階の種多様性は自然の推移の中で極相に達するにつれて減少していきますが、人間のマネジメントで様々な生態系を共存させ高い生物多様性に貢献することができます。生態系の回復プロセスは人間の助力による生物多様性の向上と特に適合します60,70,71。確かに、軽くあるいは集中的に使用される二次植生が種多様性とその量において一次植生を超えると世界中で報告されています72。世界で消費される食料のカロリーの90%はわずか30種の植物から得られていると推定される中、歴史的に人類に食べられてきた植物種は30000種を超えるとされ73、これは、農地生態系の拡張に向けて利用されていない植物の遺伝資源の広大なレパートリーがあることを示しています。
もし我々がそういった生態系の拡張を食料生産の大部分(特に、発展途上国の小規模農家35,36)において達成することができれば、人新世における成果として、我々の社会-生態系システムを持続的に動かす大きな力となり得ます。図3(a1-a3)は、生態学的なレジームシフトを防ぐことや逆転することについて、可能性のあるメカニズムとシナリオを示しています。(a1)は橙の矢印で地球システムの、多種の生命を擁する複雑な生態系への進歩を表しています。灰色の矢印は歴史的にin culturaの農業によって地球の生物多様性が破壊され、自然の物質循環が損なわれ、グローバルレジームシフトが起きる閾値(Tの線で、図1のlimit of natural resilienceに相当)に向かっていることを表しています。赤の矢印はin natura生態系拡張が生態学的な複雑性を再構築する力となることを示しています。これに同方向で付随する茶色の矢印は、惑星環境を可住状態にするためのテラフォーミングを表します。
(a2)の青線は生態系のヒステリシスの概略を表します。x軸は、農地への転換や環境汚染、その他の環境破壊などの人間による影響の強さを表します。y軸は、面積当たりの生物多様性などの、生態系の状態を表します。これが参考文献1に基づいた生態学的ステートシフトの典型的な遷移図です。赤線は、人間活動がある程度生態系の状態を改善するように動いて生態系が拡張された場合の予測されるカーブを表しています。結果として青線に比べて右にシフトしています。破線の矢印は2つの状態間のレジームシフトを表しています。これは下方向の生物多様性崩壊と上方向の生物多様性回復であり、非可逆的プロセスです。
(a3)は、推測される生態系のステートシフトの防止あるいは逆転のダイナミクスを表しています。x軸は時間を表していて、2045年までに起こると予想されるグローバルステートシフト(地球生態系の全球的崩壊)の時点も示されています。y軸は(a2)と同様です。隙間のない(破線でない)線は(a2)の状態遷移を時間スケールで表しています。yの値は局所生態系の生物多様性(緑)、全球規模の生物多様性(青)、局所と全体すべてが拡張された生物多様性(赤)を表しています。破線の矢印は状態遷移を表しています。それぞれ、局所生態系の崩壊(CLE:緑の下方向への矢印)、全球生態系の崩壊(CGE:青の下方向への矢印)、拡張された生態系の局所的あるいは全球的な崩壊(CAE:赤の下方向への矢印)、局所生態系の回復(RLE:緑の上方向への矢印)、全球生態系の回復(RGE:青の上方向への矢印)、拡張された局所生態系の回復(RLAE:赤の上方向への矢印2つ)、拡張された全球生態系の回復(RGAE:赤の上方向への矢印)、です。
この図が示唆することは、拡張された生態系(赤線)が生物多様性の減少を緩和し、レジームシフトのギャップを小さくしていて(橙色の矢印)、もし食料生産の大部分においてかなりの拡張が達成された場合にはそれを防ぐことも可能であることです。
図3
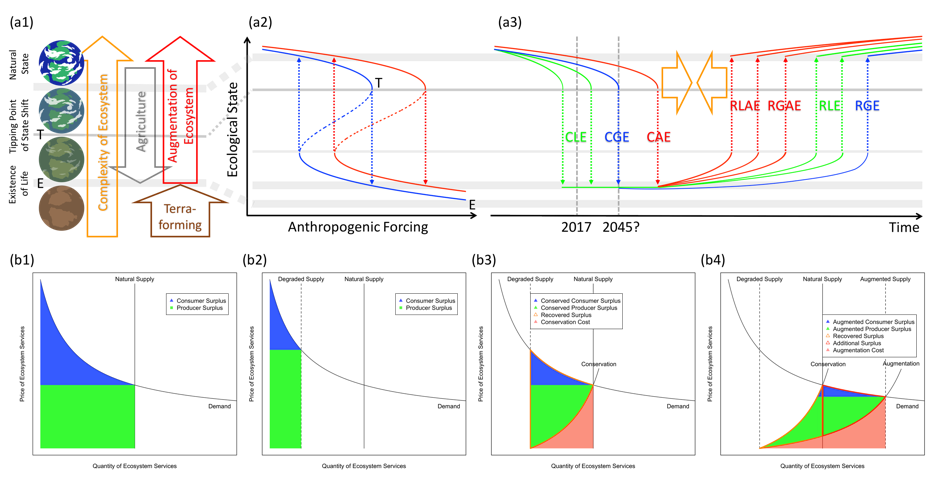
図3(b1-b4)は、開発・保全・拡張のそれぞれのシナリオについて生態系サービスの推測される収益を価格決定のメカニズムと関連して表しています。すべて生態系サービスの需要供給曲線を参考文献75に基づいて示しています。x軸は量で、y軸は価格です。
(b1)は人間による破壊がない状態での自然な供給の場合です。通常の経済学で言われるようなものとは違い、人間の需要量に関係なくコンスタントな量を供給する生態系サービスの供給曲線は垂直の閾値で表現されます。青と緑で塗られた部分はそれぞれ消費者の利益、生産者の利益を表します。
(b2)は人為的な力によって自然の供給が損なわれた場合です。供給曲線は左にシフトし、消費者、生産者の利益を小さくしています。
(b3)は人間が自然生態系を自然状態まで回復させるためにコストをかけるという典型的な保全シナリオの場合です。保全コスト(薄赤の部分)と引き換えの消費者と生産者の利益(青と緑の塗りつぶし)が((b2)に加えて)回復された利益(橙の線で区切られた部分)として予測されます。
従来の生物多様性のコンセプトでは普通、自然状態を最高の基準とし人間による破壊について議論します。それとは対照的に、生態系の拡張は種多様性操作の新たなフィールドを創り出し、実際に拡張生態系の構築やバーチャルな知識の多様性を生みます66。それに従って有用性・活用可能性のキャパシティは高まり、それが(b4)でモデル化されています。前提として、保全を超えた生態系の拡張とは自然保護状態よりも生態系サービスの供給の拡張を実現させることです。成功のケースでは、保全シナリオ(b3)に比べて追加で回復された利益(橙の線で区切られた部分)とさらに追加の利益(赤線で区切られた部分)が拡張された消費者と生産者の利益として予想され、これはより良いコスト効率を伴います(薄赤の部分)。
この、生態系サービスにおけるコストと利益の分類はグリーンエコノミー(緑の経済)における政策方針決定に基本的な市場メカニズムを供給し、補助金の分配を生態系拡張と従来の保全作業の明確な区別の下で行えることを可能にします。保全を超えた拡張は、実現された生態系の状態と管理様式によって査定可能な生態系サービスにさらなるレバレッジ効果を与えます。現状の、食や生産生態系において生物多様性をメインに据えようとする国際的なイニシアティブは、そういった生態系サービスにおける利益/コストの効率に従って(特に保全を超えた拡張augmentationを区別して)、投資を優先させるというフレームワークを組み込むべきです。適用可能性のあるプロジェクトは、次のものです。食と農地生態系の多様性を促進し局所的な適応を通して栄養状態を改善すること76、栄養に配慮した農業の発展に向けての政策決定と活動計画77、持続的な森林利用におけるカーボンオフセットと生物多様性促進の統合的市場メカニズム78、人間と動物の医学の連携による種間のヘルスイニシアティブ79、遺伝資源へのアクセスと利益配分についての名古屋議定書とそのクリアリングハウスメカニズム66,80、商業的農業と森林部門における生物多様性への正味のプラス(net positive impact)のアプローチ20、多様な発展途上国における現地での植物遺伝資源の保全などの政府主導の農地生物多様性への統合的アプローチ81、未利用や無視されてきた利用可能な生物種の持続可能な利用の促進82、多年生のメジャーな作物種の開発による炭素固定の促進や限界耕作地における低投入耕作83,84、植物種の多様性向上による多様な環境での収量と生態系サービスの適応85です。現状のイニシアティブを拡張シナリオに転換するときの基準はsupplementary material 8に示してあります。
農業以外の食の一次生産も生態系の拡張の達成を助けるポテンシャルを秘めており、産業間をうまくコーディネートすべきです。伝統的牧畜は、最も生物学的に多様なサバンナ生態系のいくつかを厳しい環境に創り出しましたが、気候変動をうまく切り抜ける為に新たな適応をする必要に迫られています86。ここ最近あらわれてきた集約放牧は土壌炭素を元の森林のレベルまで急速に固定することがわかっています87。広範囲の養殖漁業システムは副次的に水中生態系の種多様性とその量を高め88、結果として湿地における鳥類の住処にもなります89。昆虫食などの生態学的に健全な代替食の展開は森林環境の持続可能な利用において生物多様性をポジティブに刺激します28。
in natura生態系拡張(これはまさに過去のin cultura農業の歴史に比して対極にあります)の度合いが、地球システム(人間や環境の健康を含む自然資本の総体) の運命を分かつでしょう。
これらの活動を科学的エビデンスで促進するため、分野横断的なアプローチが重要となります。多様なスケールの生態学的、栄養学的ビッグデータ90,91は未来に向けて従来のシナリオの制限を克服するため、生態系拡張のより正確なダイナミクスを組み込む必要があります。栄養的リスクアセスメントを超えて、人間と生態系の統合体のシステム欠陥型の問題に対処できるシステムレベルの特性へ踏み込むべきです。個々人に最適化された医学、栄養学、縦断的オーミクスによる公衆衛生92の結合は、さらに環境の潜在的変数(図2の(a1)から(a3))を統合し、拡張された生態系サービスから得られるであろう利益に結びつけるべきです。
上記で議論されたように、食品成分の多様性と生物多様性の間の溝に橋をかけることは科学政策における喫緊の技術的課題です。持続可能性の実現には、社会-生態学的システムの回復力を築くために科学技術と社会的なサポートの協働が必要です。もっと根本的なレベルでは、公平な生産や流通形態を通して経済的不釣り合いをなくす健全で協働的な地域経済を構築することが、基本的な社会基盤と同等に重要です7。さらに廃棄物や消費、温室効果ガス排出やインフラ建設について、こうした人間の文明活動の影響が地球の限界内(planetary boundaries)に収まるように減らすあるいは抑えるべきです。これには、再生不可能な資源の節約やリサイクルのため、またエネルギー消費的なライフスタイルを環境にポジティブなものに転換するため、そして拡張された生態系サービスから得られる基本的な産物・日用品の大半を供給するために、様々なものやサービスをシェアするようにシフトした経済活動のリフォームが必要です。それに向けて、新しい取り締まりの枠組みと両立する法的システムの開発が、迅速かつ効果的な実装のために政策決定の中で必要になります。第四次産業革命において実現可能な広範囲の様式のICTも、SDGs(持続可能な開発目標)との緊密な協働の中で活用されるべきです15,66,93。技術・社会的仲介者が、自律分散型組織(DAO)の多様なステークホルダー同士のアクセシビリティと協力による相乗効果の最大化のために、オープンソースかつ全員参加型の構造に組み込まれるべきです。日本におけるソサエティ5.0のようなスマートソサエティに向けた政府のイニシアティブは、未来の自然資本のベースラインに拡張生態系を据え、全球の生物圏のポテンシャルを急成長するサイバースペースも使ってフルに活用する必要があります(図1の右上)。
先進国と発展途上国間における社会的ギャップの消失と環境負荷の差異の減少が人口転換を新たな局面に導くでしょう。特に、発展途上国における食の品質の改善と生物多様性の回復は人間開発指数(HDI)の上昇を助けますが、これは一般的に出生率の低下を招きます。同時に、HDIの高い国において出生率は再上昇することも知られています62。これは先進国と発展途上国間の出生率のギャップを埋め、年齢や地域的な人口のより平等な分布をもたらし、HDI格差を減らし、平均を高めることにつながるでしょう。HDIの要素の中で、平均余命などの健康の指標について、食料生産とそれに付随する自然環境を改革することで大幅に改善可能です。結果としてこれが人新世を、適切なスケールの人間の文明と拡張された生態系が同時に繁栄するために互いに有益な働きをする共生的地球(symbiotic Earth)へと導くことにつながり、非常にポジティブな未来を実現することができます。
終章
現在続くこの惑星の生態系破壊と付随して生じる健康リスクに対処するために、食料生産の根本的な再構築が必要です。これは単に農業における生物多様性と生産性のトレードオフの効率を良くすることや、in vitroやin vivoにおける要素的知見と、in culturaのモノカルチャー主体の既存の食料システムが前提であるコホート研究を結びつけることだけでは達成できません。むしろ、人類や他の生物の遺伝的・代謝的プランを創り出した生態系の進化の中で全球的な生命の支えとなってきたin naturaダイナミクスを組み込む必要があります。この多段階のスケールに渡る展望において、分子生物学とシステムバイオロジーであったり、フードサイエンスと医学であったり、農学や生態学と地球科学のような関連する分野を、in natura生命科学という文脈の下にICTや計算機科学のインターフェースを用いて統合、刷新するべきです。持続可能な食産業と人間の健康の高い水準を両立するため、人間の健康に関する新興の縦断的オーミクス研究は巨視的な生態学的な変数(生物多様性の破壊、保存、緩和、保全、拡張の幅に渡る)を組み入れ、区別するべきです。こうした統合には、個体の生理学から群集生態学にまで渡る異なるスケールで相互関連し文脈に応じて解釈可能な、基準となるモデルを必要とします9。
予想される社会-生態学的システムの転換点まで時間が限られており1,23,64、現場の食料生産はそれが来るのをただ待つべきではありません。代わりに、我々人類はそれぞれの地域(特に小中規模の農家・農地)で生態系を拡張させ、この惑星の物質資源の制限を考慮し長期的な視点を持ちつつその経済的・生態学的利益を最大化させるよう事前に動かねばなりません。
こうした動きを促進するため、技術への投資や補助金支援、政策立案などにおいて、低投入で生物多様性重視の小規模農家に力を貸す方向に大幅に舵を切るべきです。これがボトムアップに大多数のステークホルダーに相乗効果を生み出します。2050年に世界人口は91億人に達するといわれる中、その時にそれだけの人数を養う食料生産の大部分はローカルな小規模農家たちによって築かれたin natura拡張生態系によって実現される必要があります。これには、地球規模で分散化された生産・流通システムの中で超多様性を資源効率的にマネジメントするためICTによるサポートなどの技術的投資もなされるべきです13-15,61。
SDGsは産業や科学の革命を求めており、食料生産を、前例のない改革におけるメインの柱のひとつに据えています60,93,94。人間の能力の拡張とそれが作り出す全球的な生態系の状態が、地球システムの未来の道筋を決める重要な役割を果たすでしょう。
謝辞
「野人」大塚隆氏は有用な見識を提供してくださいました。太田耕作氏と河岡辰弥氏はリサーチアシスタントとして貢献してくれました。イラストは玉木明氏によって描画・提供していただきました。Raymond Rodriguez氏と3人の匿名のレビュアーより価値のあるコメントをいただきました。株式会社ソニーコンピュータサイエンス研究所が研究資金を出資しました。